CAM
1 / fe
1 /
|
|
ARTICULO ORIGINAL |
|
|
|
1.
Introducción |
Antoni
Balagué López
|
|
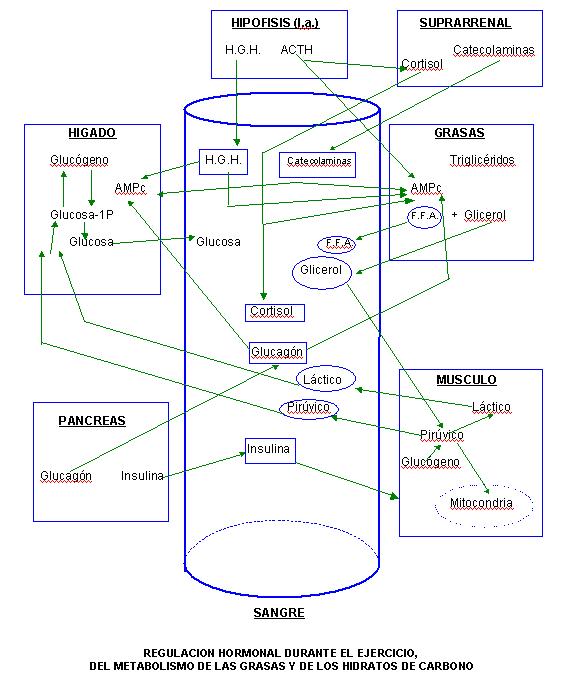
Los cambios del metabolismo lipídico e hidrocarbonado que se producen durante el ejercicio y que hemos comentado en dos artículos anteriores (I) y (2), tienen un sustrato hormonal que los desencadena y regula. Durante el esfuerzo físico se produce en el individuo humano un complejo mecanismo de movilización de determinadas hormonas que adaptan y regulan el metabolismo del individuo a la situación de esfuerzo. En este artículo sólo nos referimos a las hormonas que durante el ejercicio regulan el metabolismo de las grasas y de los hidratos de carbono: las catecolamina (adrenalina y noradrenalina), el glucagón, la insulina, la hormona de crecimiento y los glucocorticoides (cortisol).
|
|
Varios autores (3), (4), (5), (6), han encontrado un aumento de la eliminación urinaria de catecolaminas, o de uno de sus productos de degradación, el ácido vanilmandélico, durante el ejercicio. Los aumentos llegan a ser del doble con respecto a los valores basales sin que existan diferencias notables entre la producción de adrenalina y noradrenalina (7 ). Asimismo, se ha podido comprobar, que en sujetos bien entrenados se produce una menor eliminación urinaria de catecolaminas con respecto a los no entrenados (8).
Regulación Se conoce que durante el ejercicio físico existen una serie de mecanismos nerviosos que condicionan un aumento de tono vegetativo simpático. Así, estímulos nerviosos originados en el área motora de la corteza cerebral activan a las vías simpáticas al mismo tiempo que mandan impulsos nerviosos a las neuronas motoras medulares. También por vía nerviosa se activa la vía simpática a través de receptores periféricos sensibles a la bajada de la concentración de oxígeno (durante el ejercicio territorios musculares consumen notables cantidades de O2). Con el entreno esta función vegetativa simpática es menor, sin que se conozca el verdadero mecanismo de esta adaptación.
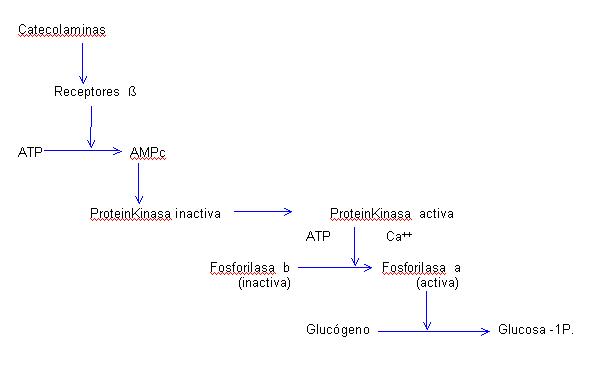
Funciones metabólicas durante el ejercicio a) Sobre los hidratos de carbono.- Las catecolaminas actlivan a la fosforilasa hepática y muscular, favoreciendo la degradación del glucógeno. La noradrenalina tiene un efecto menor que la adrenalina. Concretamente, las catecolaminas favorecen la síntesis de AMPc, a través de ATP, activando a la fosforilasa según la cadena siguiente:
|
|
|
|
|
|
Nosotros (9) hemos podido comprobar un notable aumento de los niveles en sangre de AMPc durante un esfuerzo prolongado, niveles que se mantienen elevados, a la media hora de fina lizado el esfuerzo, corroborando que el efecto glucogenolítico se debe producir a través de este nucleótido. Mediante la activación de la fosforilasa hepática se aumenta el aporte de glucosa a la sangre. Medianlte la activación de la fosforilasa muscular, se forman cantidades de piruvato que en condiciones aeróbicas es metabolizado o en condiciones anaeróbicas [1] es transformado en ácido láctico que puede volver a ser oxidado en el hígado y favorecer de nuevo la síntesis de glucógeno. [1] In 1980, by trying to clarify a terminology that was not appropriated from a biochemical point of view, Balagué et al. already published a paper (Barbany, Company and Balagué, 1980) pointing out the incorrectness and difficulty to classify the exercise in aerobic and anaerobic. However, this terms are used for practical reasons.
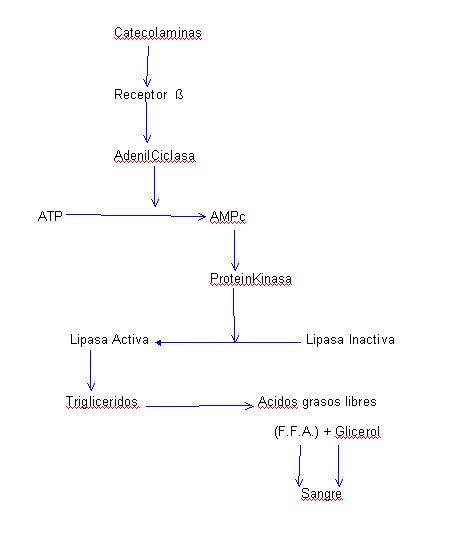
b) Sobre las grasas.- Las catecolaminas son capaces de liberar ácidos grasos a la circulación procedentes de la hidrolización de los triglicéridos almacenados en la gota de grasa de las células adiposas. Esta acción la realizan las catecolaminas activando una lipasa que está en forma inactiva pero que es convertida a forma activa por el AMPc mediante la proteinkinasa. La secuencia de cadena es la siguiente:
|
|
|
|
A través de un aumento en la secreción de catecolaminas y de AMPc podemos explicar el aumento de la concentración de ácidos grasos libres existenltes en plasma de sujetos durante el esfuerzo. La diferencia en los aumentos de la concentración de ácidos grasos libres en plasma que se encuentra en individuos entrenados y no entrenados se puede esplicar, a su vez, por la distinta capacidad de segregar catecolaminas en ambos grupos. Así en los sujetos entrenados, la liberación de catecolaminas es menor, coincidiendo con que el aumento de ácidos grasos en sangre también es menor con respecto a los no entrenados (1), (8).
|
|
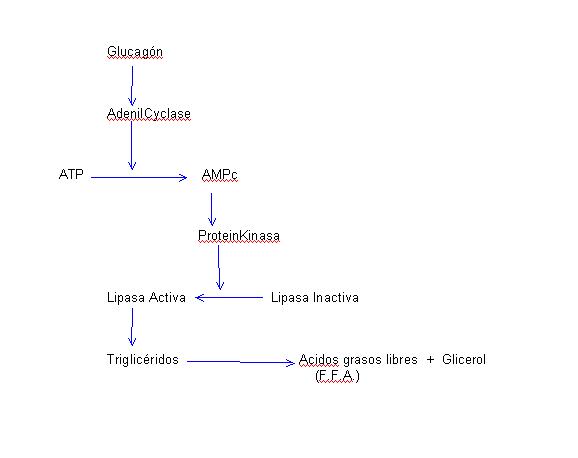
Los aumentos de glucagón durante el ejercicio físico han sido demostrados por varios autores (10), (11), (12), (13). Se ha observado que en ejercicios moderados las concentraciones sanguíneas de glucagón sólo aumentan en un 35 % mientras que en ejercicios intensos y prolongados estos aumentos pueden ser del orden de un 300 % (11 ). Asimismo, se ha podido comprobar un comportamiento completamente distinto en los niveles plasmáticos de glucagón durante el ejercicio según que los sujetos estén muy entrenados o desentrenados. En los sujetos muy entrenados incluso llega a producirse una disminución de los niveles de glucagón sanguíneo durante el ejercicio, para una vez finalizado el ejercicio, volver estos valores a sus concentraciones normales. Por el contrario, en sujetos no entrenados se produce un manifiesto aumento de los niveles de glucagón en sangre, permaneciendo todavía altos, transcurridas dos horas de finalizado el ejercicio (13 ) . Regulación Los mecanismos que controlan la secreción de glucagón durante el ejercicio físico son: 1. Activadores El aumento de tono simpático que se produce durante el ejercicio, por vía b-adrenérgica estimula la secreción de glucagón. Así un bloqueante de receptores-b aplicado durante el ejercicio, frena los aumentos de secreción de glucagón (12). Hay autores (11) que han encontrado una correlación entre los niveles de adrenaIina y noradrenalina con el glucagón en el ejercicio intenso, aunque esta correlación sólo se mantiene con la adrenalina en el ejercicio moderado. Este efecto adrenérgico probablemente se media a través de concentraciones de AMPc. Se ha podido comprobar que cuando se perfunde un páncreas aislado con concentraciones crecientes de AMPc, aumenta la secreción de glucagón (14 ). Aquí queda de nuevo resaltado el interés de los aumentos de la concentración de AMPc durante el esfuerzo (9). La bajada en la concentración sanguínea de glucosa, es un potente mecanismo de secreción de glucagón. Pero estas bajadas en la concentración sanguínea de glucosa sólo se producen después de un ejercicio intenso (2). Y sólo en este caso tendrá interés la bajada de concentración de glucosa como estímulo segregador de glucagón. El aumento de concentración de aminoácidos en sangre ocasiona un aumento de secreción de glucagón. No parece ser un efecto interesante durante el ejercicio. 2. Inhibidoras El aumento de ácidos grasos libres en sangre, crea un mecanismo de frenado sobre la secreción de glucagón. Ya hemos comentado en el trabajo anterior (1) el aumento de ácidos grasos producidos durante el ejercicio. De ahí cabe atribuir un control negativo por parte de estos ácidos grasos, en relación con el grado de entreno del individuo. Los aumentos de la concentración de glucagón durante el ejercicio nos indican el predominio de los efectos activadores sobre los inhibidores. Funciones metabólicas durante el ejercicio a) Sobre los hidratos de carbono.- El glucagón es glucogenolítico y gluconeogenético. Glucogenolítico.-El glucagón estimula la adenilcilasa de las células hepáticas. Esto conduce a la activación de la fosforilasa, y por lo tanto al incremento de la demolición de glucógeno. El glucagón no provoca glucogenólisis en el músculo. En el hígado aumenta la neoglucogénesis a partir de los aminoácidos disponibles. b) Sobre los lípídos.- La lipasa intracelular del tejido adiposo, que transforma los triglicéridos en ácidos grasos libres y glicerol, también es sensible a la acción del glucagón. El mecanismo por el cual esta lipasa es activada es semejante al comentado anteriormente:
|
|
|
|
|
|
Este efecto lipolítico del glucagón también interviene por consiguiente en el aumento de ácidos grasos libres que se producen durante el ejercicio. Existe también en este caso un paralelismo entre el grado de entreno y los niveles sanguíneos de glucagón y ácidos grasos libres.
|
|
Algunos autores (15 ), (16 ), (17 ), demuestran una disminución de las concentraciones plasmáticas de insulina durante el esfuerzo. En un trabajo reciente (13), se comprueba un comportamiento distinto en los niveles de insulina plasmática durante el esfuerzo según que los individuos estén o no entrenados. En los sujetos entrenados, en los primeros tres minutos de un esfuerzo en un cicloergómetro, se produce una subida de los valores de insulina plasmática, para ir descendiendo y al final del esfuerzo encontrarse por debajo de los valores basales. Dos horas después de haber finaJizado el esfuerzo, estos valores siguen por debajo de los valores preesfuerzo. En cambio, en los sujetos no entrenados, se produce ya durante el primer minuto de iniciado el ejercicio una subida de los niveles de insulina plasmática, niveles que se mantienen elevados a lo largo de todo el esfuerzo e incluso a las 2 horas de haber finalizado el ejercicio. Este último hallazgo concuerda con otros autores (18) que han encontrado un aumento de los niveles plasmáticos de insulina durante el esfuerzo físico en individuos no entrenados. Asimismo, las concentraciones plasmáticas de insulina durante un esfuerzo corto varían en relación a que los individuos entrenados ingieran antes de iniciar el esfuerzo una disolución rica en hidratos de carbonos (19). En los individuos que ingieren agua sin hidratos de carbono (placebo), la insulina plasmática cae durante el ejercicio por debajo de los valores basales, mientras que en los sujetos que han ingerido hidratos de carbono, aunque durante el ejercicio se produce una disminución de la insulina plasmática, ésta no cae nunca por debajo de los valores basales. Esta experiencia sugiere una modificación de la eficacia de los estímulos que durante el esfuerzo se encargan de variar la secreción de insulina, relacionada con los niveles de glucosa en sangre. Regulación La secreción de insulina pancreática durante el esfuerzo está regulada por: Los niveles de la concentración de glucosa en sangre.- Este no parece ser un mecanismo importante de regulación durante el ejercicio físico, puesto que los niveles plasmáticos de glucosa varían poco. Tampoco este mecanismo de regulación puede tener interés en ejercicios intensos y prolongados, cuando baje la concentración de glucosa sanguínea creando un estímulo frenador de la secreción de insulina. Los niveles plasmáticos de glucagón.- El glucagón es una hormona estimulante de la secreción de insulina por una acción directa sobre las células beta y por una acción indirecta a través de los aumentos de glucosa en sangre. Durante el ejercicio físico, cabe destacar que los aumentos de glucagón crean un efecto directo activador de la secreción de insulina. La concentración sanguínea de catecolaminas.- Los aumentos en sangre de noradrenalina inhiben la secreción de insulina (20), (21). Los aumentos de secreción de catecolaminas que se producen durante el ejercicio, actuarán como un importante mecanismo frenador de la secreción pancreática de insulina. Los niveles sanguíneos de AMPc.- Los aumentos en sangre de AMPc estimulan la secreción de insulina, y ya hemos comentado previamente que el AMPc aumenta su concentración sanguínea durante el ejercicio. La somatostatina.- Los aumentos de somatostatina frenan la secreción de insulina. Durante el ejercicio existe un aumento de la secreción de hormona de crecimiento, frenándose muy probabJemente la liberación de somatostatina. En resumen pues, durante el ejercicio físico, existen varios mecanismos excitadores y frenadores de la secreción de insulina que actúan simultáneamente. Este hecho permite explicar los hallazgos de distintos autores, en parte contradictorios, sobre las modificaciones plasmáticas de insulina durante el esfuerzo especialmente al considerar las variaciones de secreciones hormonales en relación con el grado de entreno del individuo.
Funciones metabólicas durante el ejercicio a) Sobre el metabolisnto de los hidratos de carbono.- El aumento de la concentración plasmática de insulina crea: Aumento de la entrada de glucosa al músculo y a otros tejidos por una acción sobre la membrana celular. Aumento de la entrada de glucosa al tejido adiposo. Aumento de la síntesis de glucógeno muscular. Disminución del AMPc, de la gluconeogénesis y de la síntesis de glucógeno en el hígado. El efecto importante que la insulina pudiera tener sobre la captación muscular de glucosa durante el ejercicio físico, queda en duda por las oscilaciones (aumentos o descensos) de los niveles sanguíneos de insulina plasmática que se producen durante el esfuerzo. Así como, por el hecho comprobado de que la penetración de glucosa en el músculo esquelético en ejercicio está aumentada aún en ausencia de insulina (27). b) Sobre el metabolismo de los lípidos.- El aumento de la concentración plasmática de insulina crea: Aumento de la síntesis de ácidos grasos en el tejido adiposo. Aumento de la síntesis de fosfato de glicerol en el tejido adiposo. Aumento del depósito de triglicéridos en el tejido adiposo. Aumento de la síntesis de lípidos a nivel del hígado. Las dudas sobre la importancia fisiológica de la insulina durante el ejercicio físico han sido comentadas en el párrafo anterior.
|
|
5. Hormona de Crecimiento (HGH) Es bien conocido que durante el ejercicio se producen elevaciones de la concentración plasmática de hormona de crecimiento (22), (23), (24), (25). Incluso se ha utilizado una prueba de ejercicio en un cicloergómetro como un « test » clínico para la exploración de la liberación hipofisaria de hormona de crecimiento (26). Los niveles plasmáticos de hormona de crecimiento durante el esfuerzo, varían con el entreno. En sujetos bien entrenados, los aumentos sanguíneos de HGH producidos con el ejercicio son menos marcados que en los no entrenados. A su vez una vez finalizado el ejercicio, los niveles elevados de hormona se prolongan durante más tiempo en los sujetos no entrenados que en los sujetos bien entrenados (25). Regulación Los mecanismos que determinan el aumento de hormona de crecimiento durante el ejercicio son desconocidos. No obstante cabe plantear: Que la hipoglucemia estimula la secreción. Este no parece ser el mecanismo que actúe normalmente durante el ejercicio por idénticas razones a las comentadas anteriormente. Que el glucagón estimula la secreción de HGH, y durante el ejercicio físico, tal como hemos comentado, aumentan notablemente los niveles plasmátieos de glucagón. Que puede existir un efecto directo desde el área motora sobre el hipotálamo, gobernando la secreción de dicha hormona por control nervioso. Que los niveles altos de lactato en sangre, estimulan la secreción de HGH. Así, cuando se administra endovenosamente lactato, se produce un aumento significativo de la concentración plasmática de HGH (25). Esta experiencia permitiría explicar una coordinación entre la recuperación a valores normales de los niveles plasmáticos de lactato y de HGH al finalizar el esfuerzo, tanto más velozmente cuanto mayor es el grado de entreno del sujeto. Realmente debemos preguntarnos si los niveles sanguíneos de lactato pueden ser un factor importante en la secreción de HGH durante el esfuerzo. Que los niveles de cortisol y ácidos grasos libres en sangre frenan la secreción de HGH. Durante el ejercicio físico el cortisol y los ácidos grasoso libres están aumentados. En definitiva pues existen, al igual que para otras hormonas, efectos excitantes e inhibidores de la secreción de HGH durante el ejercicio físico que, de alguna manera, controlan los aumentos de HGH que se producen durante el esfuerzo. Funciones metabólicas durante el ejercicio a) Sobre el metabolismo de los hidratos de carbono.- En una fase inicial del ejercicio, la HGH aumenta la capacidad de captación muscular de glucosa. En una fase más posterior, se inhibe la capacidad de captación muscular de glucosa y se aumenta la producción de glucosa a partir del glucógeno hepático. La acción glucogenolítica hepática, se atribuye a una acción por parte de la HGH favorecedora de la síntesis de determinadas proteínas necesarias para la síntesis de AMPc (28). b) Sobre el metabolismo lipídico.- En una fase inicial la HGH favorece la captación muscular de ácidos grasos (28). En una fase más tardía favorece la lipólisis, a través de la síntesis de AMPc. Esta hormona puede ser junto con las catecolaminas y el glucagón un efecto importante sobre el aumento de ácidos grasos libres que se producen en sangre durante eJ ejercicio.
|
|
Durante el ejercicio físico se producen notables aumentos de los niveles de cortisol sanguíneo, siendo estos aumentos tanto más marcados cuando mayor sea la intensidad del esfuerzo físico. Con el entreno, los aumentos del cortisol plasmático son menos manifiestos (29). Los cambios del cortisol se originan a nivel central, variando con el esfuerzo los niveles de ACTH. Se ha comprobado que un ejercicio físico de actividad máxima induce un aumento de la secreción de ACTH hipofisaria (30). Por otro lado, talmbién se ha comprobado que la respuesta de la corteza suprarrenal a la administración endovenosa de una dosis de ACTH, es la misma en individuos entrenados que en los no entrenados (31). Regulación Es difícil en la actualidad aclarar cuáles son los mecanismos que contribuyen a incrementar las secreciones de ACTH y glucocorticoides durante el ejercicio. Nos podríamos refugiar cómodamente en el concepto de «stress», atribuyendo al ejercicio físico una función estresante para el organismo y desencadenante de la secreción de ACTH y cortisol. No obstante hay que pensar en algún mecanismo más fisiológico y así no podemos olvidar que si la corteza cerebral, concretamente en sus funciones motoras, es capaz de activar a los centros reguladores de la actividad cardíaca, del tono vasomotor y de la respiración, también pudiera de alguna manera activar la secreción hipotalámica de RF- ACTH y de esta manera, gobernar la función glucocorticoide. Funciones metabólicas a) Sobre los hidratos de carbono.- Es clásicamente conocido que con la administración de glucocorticoides se produce una elevación de los niveles de glucosa en sangre con un aumento del contenido de glucógeno hepático y muscular. Estos efectos son debidos a un efecto neoglucogénico a expensas de aminoácidos, lactato y piruvato fundamentalmente. El efecto neoglucogénico a expensas de las grasas no parece ser tan importante en la regulación de los glucocorticoides. b) Sobre las grasas.- La ACTH tiene un efecto lipolítico, activando a la adenilciclasa que a su vez activa a la lipasa de las células grasas. Los glucocorticoides también incrementan la actividad de la lipasa, aunque es a través de un proceso más lento que requiere de una síntesis de una nueva proteína y es independiente de los efectos del AMPc.
|
|
|
|
|
|
|
|
Hormone:
{G. hormòn, pres. part. of hormaõ, to rouse or set in motion}. A
chemical substance, formed in one organ or part of the body and carried
in the blood to another organ or part; depending on the specificity of
their effects, h.’s can alter the functional activity and sometimes
the structure, of just one organ or of various number of them. Catecholamines:
Pyrocatechols with an alkylamine side chain. Examples of biochemical
interest are epinephrine, norepinephrine and dopa. Dopa:
3,4- dihydroxyphenylalanine; an intermediate in the catabolism of
phenylalanine and tyrosine and in the biosynthesis of norepinephrine,
epinephrine and melanin; the L form, levodopa, is biologically active. Epinephrine=Adrenaline:
3,4-dihydroxy-a
{methylaminomethyl} benzyl alcohol; a catecholamine that is the chief
neurohormone of the adrenal medulla of most species. It is the most potent
stimulant (sympathomimetic) of adrenergic a
and b
receptors. Norepinephrine=Noradrenaline:
Levarterenol; 1-a-(aminomethyl)-3-4-dihydroxybenzyl
alcohol; a catecholamine hormone of which the natural form is D,
although the L form has some activity; the base is considered to be the
postganglionic adrenergic mediator. It is present in the adrenal medulla
and in adult animals of most species in much smaller amounts than is
epinephrine; possesses the excitatory actions of epinephrine, but has
minimal inhibitory effects. Glucagon:
Hyperglycemic-glycogenolitic factor; HG factor; pancreatic hyperglycemic
hormone; a hormone consisting of a straight-chain polypeptide of 29
residues (bovine g.) extracted from pancreatic alpha cells. Insulin:
A peptide hormone, secreted by beta cells in the islets of Langerhans,
that promotes glucose utilization, protein synthesis and the formation
and storage of neutral lipids; obtained from various animals and
available in a variety of preparations. Somatostatin:
Somatotropin release-inhibiting factor; a tetradecapeptide capable of
inhibiting the release of somatotropin by the anterior lobe of the
pituitary gland. Growth
Hormone (GH)=Somatotropin: Growth or pituitary growth hormone;
somatotropic hormone; a protein hormone of the anterior lobe of the
pituitary, produced by the acidophil cells, that promotes body growth,
fat mobilization and inhibition of glucose utilization; diabetogenic
when present in excess. Glucocorticoid:
Glycocorticoid. Any steroid-like compound capable of significantly
influencing intermediary metabolism such as promotion of hepatic
glycogen deposition and of exerting a clinically useful
anti-inflammatory effect. Cortisol is the most potent of the naturally
occurring g.’s; most semisynthetic g.’s are cortisol derivates. Cortisol:
Hydrocortisone. Cortisone:
{former acronym for corticosterone}. 17a,
21-dihydroxy-4-pregnene-3, 11,20-trione; 17a-hydroxy-11-dehydrocorticosterone;
a glucocorticoid not normally secreted in significant quantities by the
human adrenal cortex. ACTH=Adrenocorticotropic
Hormone: corticotropin; adrenocorticotropin; adrenotropin;
corticotropic or adrenotropic h.; the h. of the anterior lobe of the
hypophysis which governs the nutrition and growth of the adrenal cortex,
stimulates it to functional activity and also possesses extraadrenal
adipokinetic activity; it is a polypeptide containing 39 aminoacids, but
exact structure varies from one species to another; sometimes prefixed
by a
to distinguish it from b-corticotropin.
|
|
|
|
ADAM, K., NOWACKI, P. und SCHMID, E. (1968). Untersuchugen über die Sympathico-Adrenale Reaktion bei Hochleistungsportlern im Training und im Wettkampf. Sportartz v. Sport Med., 19, 389. BARBANY, J.R., BALAGUÉ, A. y COMPANY, X. (1978). Metabolismo de los Carbohidratos en el Ejercicio. Apuntes de Medicina Deportiva, 60 (15). BOTTGER, I. SCHLEIN, E. FALOONA, G.R., KNOCHEL,J.P. and UNGER,R.H. (1972). The Effect of Exercise on Glucagon Secretion. J. Clin. Endocrinol. Metab., 35, 117. BUCKLER, J. (1971). Growth Hormone Levels with Exercise. Arch. Dis. Child., 46, 399. COMPANY, X., BALAGUÉ, A. y BALLESTA, A. (1978). Algunas Modificaciones de la Mioglobina Sérica en un Grupo de Nadadores Durante el Esfuerzo. Comunicación presentada al I Congreso Nacional de la Sociedad Española de Química Clínica. Barcelona, octubre 1978. COMPANY, X., BARBANY,J.R. y BALAGUÉ, A (1978). Metabolismo Lipídico en el Ejercicio. Apuntes de Medicina Deportiva, 59 (15). BARBANY, J.R., COMPANY, X. y BALAGUÉ, A. (1980). Bases Bioquímicas del Trabajo Muscular Anaeróbico. Apuntes de Medicina Deportiva, 65 (17), 9-14. BUCKLER, J.M.H. (1972). Exercise as a Screening Test for Growth Hormone Release. Acta Endocrinologica, 69, 219. FORTUNO, G., FRAIOLI, F. and ISIPORI, A. Valutazione della Risposta Incretora Ipofisaria all’Activitá Fisica. FRENKL, R., CSALAY, L., SCAKRAR, G. und LANGFY, G. (1971). Untersuchung der ACTH-Wirkung auf den Steroidspiegel des Plasmas im Trainerten und im Untrainierten Organismus. Med. und Sport, 4, 122. GALBO, H., HOLST, J.J. and CHRISTENSEN, W.J. (1975). Glucagon and Plasma Catecholamine Response to Graded and Prolonged Exercise in Man. Journal of Applied Physiology, 38, 70. GANONG, W.F. (1976). Manual de Fisiología Médica. Edt. El Manual Moderno, página 294. GERBERGER, G., KEIBEL, D., LANGER, H. und PICKENHAIN, L. (1975). Steverungssebenen Molekulare Mechanismen und Dynamik der Hormonellen Regulation im Menschlichen rganismus. Med. v. Sport, 15, 97. HELLEMANS, J. (1978). Het Energie Metabolisme bij Getrainde en Ongetrainde individuen. Gennees-Kunde en Sport, 6, 130. HOFFMANN, H.D., FIEDLER, H. und KIBITTEL, W. (1972). Der Einfluss von Fahrradergometerait Sparring und Wettkampt auf die Plasmainsulin Konzentration und den Blutzuckerspiegel bei Boxern. Med. und Sport, 4, 119. KARAM, J.H., GRASSO, S.G., WEGIENKA, L.C., FRODSKY, G.M. and FORSHAM, J.H. (1966). Effect of Selected Hexoses, of Epinephrine and of Glucagon on Insulin Secretion in Man. Diabetes, 15, 571. KEIBEL, D. (1974). Nebemniereninden-Hormone und Sport Liche Leistung. Med. und Sport, 65 (3). MARLISS, E.B., GIRARDIER, L., SEYDOUX, J., WOLLHEIM, C.B., KANAZAWA, Y., ORCI, L., RENOLD, A.E. and PORTE, D. (1973). Glucagon Release Included by Pancreatic Nerve Stimulation in the Dog. J. Clin. Invest., 52, 1246. METZE, R., und LINKE, P.G. (1971). Adrenalin und Noradrenalin Ausscheidung bei Fußballspielern. Mediz. v. Sport, 23 (1). METZE, R., LINKE, P.G. und MANTEI, E. (1971). Der Katekolaminumsatz bei Trainierten und Untrainierten Jugendlichen und Erwachsenen. Med. v. Sport, 11, 327. NOWACKI, P. (1969). The Turnover of Sympathico Adrenal Hormones of Sportsmen in Training Anticipation and During Competition Judged by Measurements of the Urinary Excretion of 3-metnoxy 4-hydroxy Mandelic Acid. Biochemistry of Exercise. Medicine and Sport, 3, 205. Karger: Basel. ORARA, S., VAPAATALO, H., SAARELA, J. and REINILA, M. (1974). Blood Glucose, Serum F.F.A. and Serum Insulin Levels After the Intake of the Carbohydrate Rich Solution Before Exercise in Man. The Journal of Sports Medicine and Physical Fitness, 73 (14) POP, T., CIOARA, Z., DRAGAN, J. and VTA, I. (1975). Recherches Concernant les Variations de l´Hormone de Croissance Plasmatique chez les Esportifs de Haute Competition sans l’Influence au stress Psychique. Medicine du Sport, 38 (1). PORTE, D., GRABER, A., KUZUYA, T. and WILLIAMS, R.H. (1966). The Effect of Epinephrine on Immunoreative Insuline Levels in Man. J. Clin. Invest., 45, 228. PRUET, E.D.R. (1970). Plasma Insulin Concentrations During Prolonged Work at Near Maximal Oxygen Uptake.Journal of Applied Physiology, 29, 155. ROTH, J., GLICK, S.M., YALOW, R.S. and BERSON, S.A. (1963). Secretion of Human Growth Hormone: Physiologyc and Experimental Modification. Metabolism, 12, 577. ROTTINI, E., COZZOLINO, G., DOMINICI, G. and D’OVIDIO, M. (1971). Physical Activity and Blood Insulin, Sugar and NEFA Valves in Normal Untrained Subjects. The Journal of Sports Medicine and Physical Fitness, 1 (1). SARVIHARJV, P.J. (1973). Effect of Physical Exercise on the Urinary Excretion of Catecholamines and 17-hidroxycorticoteroides in Young Healthy Men. The Journal of Sports Medicine and Physical Fitness, 14, 171. SUTON, J.B., YOUNG, J.D., LAZARUS, L., HICKIE, J.B. and MARKUYTIS, J. (1960). The Hormonal Response to Physicl Exercise. Austr. Ann. Med., 84 (18). TCHIBITCHIAN, D.A. (1969). Sostoinanie Simpato-Adrenalovoi Sistemy ou Iounykg Legkoatletov pri Fizitecheskoi Nagrouzke na Vynoslivost. Teor. Prakt. Fiz. Kovlt, 2, (32). YOUNG, F.G. (1986). Growth Hormone. Edit. A, Pecile and E.E. Muller. Excerpta Médica: Amsterdam, pág. 139. WEIR, G.C., KNOWTON, S.D. and MARTIN, D.B. (1975). Nucleotide and Nucleoside Stimulation of Glucagon Secretion. Endocrinology, 97, 932.
|
|
|
| © 1979 Antoni Balagué López TODOS LOS DERECHOS RESERVADOS |
 © 2003 motricidadhumana.com |